Российская академия наук
Уральское отделение
Коми научный центр
Научные доклады
Выпуск 389
Г.А.Ивашевский
ЗООГЕОГРАФИЧЕСКИЙ АНАЛИЗ ПАРАЗИТОВ РЫБ БАССЕЙНА СЕВЕРНОЙ ДВИНЫ
Доклад на заседании
президиума
Коми научного центра
УрО Российской академии наук
Сыктывкар 1997
УДК 576.89 055(02)7
Г.А.Ивашевский. Зоогеографический анализ
паразитов рыб бассейна Северной Двины. - Сыктывкар,
1997. - 21 с. (Научные доклады / Коми научный центр УрО Российской академии
наук; Вып. 389).
Обобщены данные по видовому
составу, географическому
распространению и биотопическому распределению ихтиопаразитов бассейна Сев.
Двины. Основное внимание уделено разделению бассейна на участки с
относительно однородной фауной паразитов: наибольшее сходство имеют бассейны
верхней и нижней Сухоны, несколько ниже их сходство с р. Вычегда, оз.Кубенское
и р.Юг. Показана историческая обусловленность такого разделения, а также
возможные пути формирования фауны, что нашло отражение в распределении
понто-каспийских видов по трем выделам: 1) р.Сухона; 2)р.Вычегда; 3) р.Юг.
Подчеркивается важность учета экологических различий биотопов для зоогеографического
анализа: бореальные предгорные виды сосредоточены в малых реках, паразиты
карася и красноперки - в озерах.
G.A.Ivashevskiy. Geographical
analisis of fauna of fish parasites of the Northern Dvina drainage-basin. -
Syktyvkar, 1997. - Pp. 21 (Scientific Reports / Komi Science Centre, the Urals
Division, the Russian Academy of Sciences; Issue 389).
The results of investigations of fauna parasites, their geographycal and
ecologycal distribution in the Northern Dvina drainage-basin are generalized.
The drainage-basin is divided in to some divisions with homogeneous fauna
parasites: the greatest similarity is marked between the Upper Suhona and the
Lower Suhona rivers, lower their similarity with the Vichegda river, the
Kubenskoe lake and the Yug river. Historical causes this division and possible
ways of fauna formation are presented, that is reflected in distribution of
Black Sea - Caspian Sea species on three divisions: 1)the Suchona river; 2)the
Vichegda river; 3)the Yug river. Importance of ecological differences
reservoirs for geographical analisis of fauna is underlined: borealis
pre-mountainous parasites species inhabit
small rivers, parasites of crucian and rudd (redeye) inhabit lakes.
Редакционная коллегия
М.П.Рощевский (отв.редактор), Б.А.Голдин (зам.отв.редактора), В.М.Ануфриев (отв.секретарь), А.Е.Ванеев, В.А.Головко, Н.А.Громов, А.В.Кучин, В.Н.Лаженцев, Н.А.Манов, А.Д.Напалков, Ю.С.Оводов, В.П. Подоплелов, Э.А.Савельева, А.Ф.Сметанин, А.И.Таскаев, Н.И.Тимонин, М.В.Фишман, Н.П.Юшкин
© Г.А.Ивашевский, 1997
© Коми научный центр УрО Российской АН, 1997
Важность паразитологических
данных для зоогеографии обусловлена тем, что паразиты рыб - это, с точки зрения
систематики, крайне разнородная группа, связанная с разнообразным комплексом хозяев, включающим представителей
почти всех классов многоклеточных животных пресноводных водоемов. Исследования
зоогеографии паразитов рыб СССР начаты еще В.А.Догелем (1947). Далее, с
накоплением информации по фаунистике паразитов продолжаются и зоогеографические
изыскания. В частности, изучению паразитофауны рыб Европейского округа Ледовитоморской
провинции посвящены работы С.С.Шульмана (1958), И.В.Екимовой (1976). Работы
Г.Н.Доровских (1990) и О.Н.Пугачева (1990) опубликованы недавно и дополняют
друг друга: в первой из них рассматривается бассейн Сев.Двины, во второй -
крупные водоемы Ледовитоморской провинции, но без учета Сев.Двины. Работа
Г.Н.Доровских (1990) по зоогеографии паразитов рыб бас.Сев.Двины в настоящее
время является не полной, так как построена на анализе распространения только
149-ти видов паразитов (вместо известных сейчас 189-ти видов).
Целью дальнейшей работы было
провести зоогеографический анализ и рассмотреть формирование паразитофауны рыб
бас. Сев.Двины с современных позиций. Кроме того, стояла задача проследить
характер распределения ихтиопаразитов по бас. Сев.Двины и определить его возможные
причины.
В настоящее время из литературных источников для бас.Сев.Двины
известно 177 видов паразитов (Гнедина,
Савина, 1930; Шульман, Шульман-Альбова, 1953; Кудрявцева, 1957, 1959, 1962,
1966; Шульман, 1958; Хотеновский, 1985;
Доровских, 1988; Ивашевский, Доровских, 1993; Ивашевский, 1994; 1995; 1996), а
также, по впервые публикуемым сведениям в бас.Сев.Двины имеется еще 12
следующих видов (определение автора):
Myxobolus cybinae Mitenev, 1971; Myxobolus bliccae Donec et Tozyjakova, sp.nov.: in Donec et Schulman,1984; Myxobolus albovae Krassilnikova
in: Schulman, 1966; Myxobolus guyenoti Naville, 1928; Myxobolus magnus Awerinzew,
1913; Trichodina nemachili Lom, 1960; Trichodina pediculus Ehrenberg, 1838; Trichodina acuta Lom, 1961; Dactylogyrus
yinwenyingae Gussev, 1962; Diplostomum
petromyzifluviatilis Diesing,
1860;
Diplostomum helveticum Dubois,1929; Desmidocercella
sp.
В последние годы значительно
увеличилось число известных для рассматриваемого бассейна паразитических видов
(40 видов), что связано с географическим расширением района исследований (реки
Юг, Луза); с увеличением разнообразия
изучаемых биотопов за счет малых рек; а также за счет паразитов впервые
исследованных видов рыб и круглоротых (минога, голец усатый) и уточнению
видового состава паразитов изучавшихся ранее хозяев (красноперка, золотистый
карась, речной гольян, молодь разных видов).
Для анализа сходства ихтиопаразитофаун водоемов проведено их
сравнение по фаунистическим комплексам и по видовым спискам. В последнем случае
в качестве показателя соответствия выбран индекс Чекановского-Съеренсена.
Классификация фаун проведена методом кластерного анализа по принципу одиночного
присоединения (Песенко, 1982).
ЗООГЕОГРАФИЧЕСКИЙ АНАЛИЗ ИХТИОПАРАЗИТОВ БАССЕЙНА
СЕВЕРНОЙ ДВИНЫ
Поскольку зоогеографический анализ
ихтиопаразитофауны Ледовитоморской провинции (Пугачев,1990 и др.) и ее
европейской части (Доровских,1990 и др.) уже неоднократно проводился, основное
внимание в работе уделим характеру географического распространения и формированию
паразитофауны рыб в пределах бас.Сев.Двины.
По имеющимся ихтиопаразитологическим данным, ссылкам, которые
приведены выше, бас.Сев.Двины в настоящее время можно разбить на пять участков
(выделов) - это бассейны: 1) оз.Кубенского, 2) верхней Сухоны (сюда же на
основании палеогеографических и паразитологических данных относим среднее
течение р.Сухона), 3) нижней Сухоны, 4)р.Юг (с р.Луза) и 5) средней Вычегды.
Следовательно, при разграничении бас.Сев.Двины на выделы впервые рассмотрено
распределение ихтиопаразитофауны на всей территории бассейна, за исключением
его нижнего течения (данные по которому фрагментарны). При сравнении паразитофаун
бас.Сев.Двины и прилежащих бассейнов учтены материалы по бас.Сев.Двины в целом.
Фаунистические комплексы в паразитофауне рыб
бассейна Северной Двины
Паразитофауна рыб бас.Сев.Двины состоит, в основном, из представителей
трех фаунистических комплексов: бореального равнинного, арктического
пресноводного (АП) и бореального предгорного (БП). Бореальный равнинный
комплекс подразделяется на две группы: понто-каспийские (ПК) - виды южного
происхождения и собственно бореальные равнинные (БР) - северного происхождения
(Шульман, 1958). В пределах бореального равнинного фаунистического комплекса
выделяют еще группу верхнетретичных видов, не оспаривая ее самостоятельности,
мы условно объединили ее с ПК видами, поскольку обе группы имеют южное происхождение,
что дает возможность объединить их при анализе истории формирования
паразитофауны бассейна, полностью расположенного на территории, подвергавшейся
оледенению. Принадлежность видов к тому или иному фаунистическому комплексу
определяли по литературным данным (Гусев, 1955; Богданова, Никольская, 1965;
Пугачев, 1983 а, б ; Определитель..., 1984, 1985, 1987; Митенев, 1986, 1988 и
др.). Из общего количества ихтиопаразитов бас.Сев.Двины (189 видов) представители
бореального равнинного комплекса составляют 82.6% , из них группа БР - 55.6% ,
ПК - 27.0% , видов АП комплекса - 9.0% , БП - 6.9%, виды солоновато-водного комплекса
составляют ничтожную долю и далее не учитываются (рис.1).
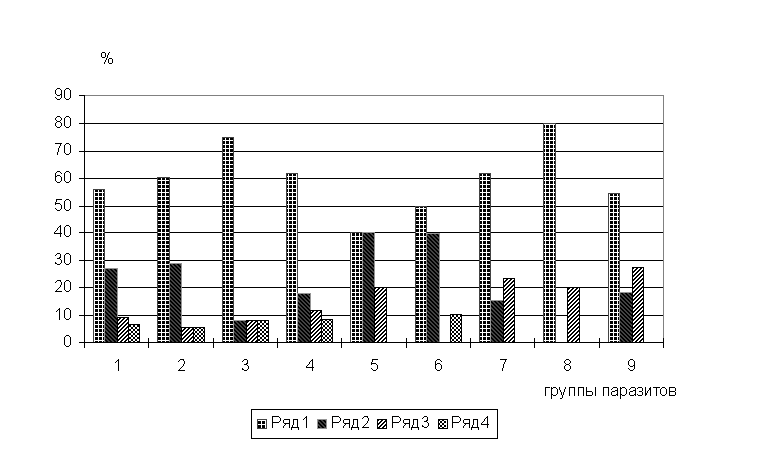
Рис.1. Соотношение доли фаунистических комплексов в паразитофауне рыб бас. Сев.Двины.
По оси абсцисс: 1 - паразитофауна в целом, 2 - миксоспоридии, 3 - паразитические инфузории, 4 - трематоды, 5 - нематоды, 6 - моногенеи, 7 - цестоды, 8 - скребни, 9 - ракообразные.
Ряды: 1 - бореальные равнинные виды (БР), 2 - понто-каспийские (ПК), 3 - арктические пресноводные (АП), 4 - бореальные предгорные (БП).
Рассмотрение доли фаунистических комплексов внутри разных классов
паразитов бас.Сев.Двины дало результат, сходный с аналогичным распределением
паразитов всей Ледовитоморской провинции (Пугачев, 1990), отличия получены лишь
по трематодам и нематодам, что, тем не менее, коренным образом изменило общую
картину распределения. Из всех классов паразитов по соотношению среди них
представителей разных фаунистических комплексов выделили три группы. Первая
группа, включающая миксоспоридии, паразитические инфузории, трематоды и нематоды
характеризуется тем же распределением по фаунистическим комплексам, что и
паразитофауна в целом; вторая группа состоит только из моногеней, здесь
отсутствуют представители АП комплекса при значительной доле БП видов; в третью
группу входят: цестоды, скребни (промежуточными хозяевами которых являются
свободноживущие ракообразные) и паразитические рачки (рис.1). То есть, у всех
систематических групп паразитов, связанных с ракообразными, доля ПК видов сильно
занижена. Причиной последнего, возможно, является то, что фауна бас.Сев.Двины
(и Печоры) отличается от таковых, принадлежащих смежным территориям, прежде
всего обедненной фауной ракообразных (Зверева,1969).
Продолжая тему фаунистических комплексов, рассмотрим их доли в паразитофауне каждого выдела
(рис.2).
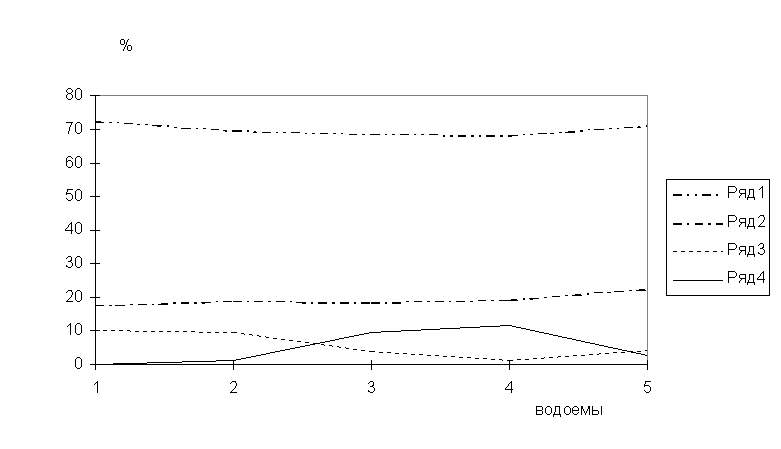
Рис.2. Соотношение доли фаунистических комплексов в паразитофауне рыб на разных участках бас. Сев.Двины.
По оси абсцисс: 1 - бас. оз.Кубенское, 2 - бас. верхней Сухоны, 3 - бас. нижней Сухоны, 4 - бас. р.Юг, 5 - бас. р.Вычегда.
Ряды: 1 - бореальные равнинные виды (БР), 2 - понто-каспийские (ПК), 3 - арктические пресноводные (АП), 4 - бореальные предгорные (БП).
Соотношение доли АП и БП видов в паразитофауне рыб на некоторых участках бас.Сев.Двины во многом
обусловлено их недостаточной изученностью, что наглядно показано на рис.2
пересечением линий, обозначающих доли АП и БП комплексов. На протяжении исследованной
части бас. Сев.Двины лидирующее положение занимают виды БР и ПК групп, доля их
в разных выделах достаточно стабильна. Постоянство доли ПК видов вызывает
сомнения в исключительности р.Вычегды как основного пути проникновения
гидробионтов в бас. Сев. Двины из бас. Волги в четвертичный период, на что
указывалось ранее (Берг, 1949; Зверева, 1969), вероятно, вследствие
неизученности фауны и истории ее формирования
рек Юг и Луза.
Обратимся к работам по палеонтологии и палеогеографии (Яковлев,
1961; Апухтин и др.,1967; Квасов, 1975), в одной из которых говорится, что “...
фауна рыб собственно Палеарктики в неогене отличалась исключительным
однообразием и имела характер ... арало-понто-каспийской ихтиофауны”(Яковлев,
1961). Формирование современной фауны бас.Сев.Двины началось после днепровского оледенения. Верхняя Сухона
представляла собой тогда Сухонское озеро со стоком в Волгу, а после отступления
валдайского оледенения сток перераспределился в Северную Двину, затем, в начале
голоцена, снова - в Волгу; все это время данное озеро было изолированным от
нижней Сухоны, долина прорыва в которую образовалась лишь на рубеже суббореального и субатлантического
времени. Вся остальная часть бас. Сев.Двины, свободная от ледника валдайского
оледенения, представляла собой Котласское озеро со стоком в бас. Волги через
русла современных рек Юг, Луза, Вычегда. Сток в северном направлении
возобновился, когда во время отступления ледников вепсовской стадии образовался
промежуток между Скандинавским и Новоземельским ледниковыми щитами. Кроме того,
имеются предположения о связи водоемов всей территории севера Восточно-Европейской
равнины, сток которых происходил в направлении Онежского озера и далее - в
Балтийское ледниковое озеро. Так же
известно о связи бас. Сев. Двины с бас.Печоры и через него - с Сибирскими реками
(Берг, 1949). Таким образом, отсутствие эндемичных для бас. Сев.Двины видов
паразитов связано с тем, что фауна данного бассейна заново сформировалась после
днепровского оледенения за счет окружающих водоемов; прежде всего за счет бас.
Волги, с которым имелись наиболее тесные контакты. Подтверждением последнего
является то, что паразитофауна бас.Сев.Двины из числа окружающих водоемов имеет
наибольшее сходство с паразитофауной бас. Волги (Доровских, 1990).
Сухона вместе с оз.Кубенским, характеризующаяся отдельной от
остальной части бас. Сев.Двины историей формирования, выделяется в
паразитологическом отношении лишь по показателю сходства фауны ПК видов; БР и
АП виды по степени сходства разных выделов приблизительно соответствуют
таковому паразитофауны в целом (рис. 3a-e).
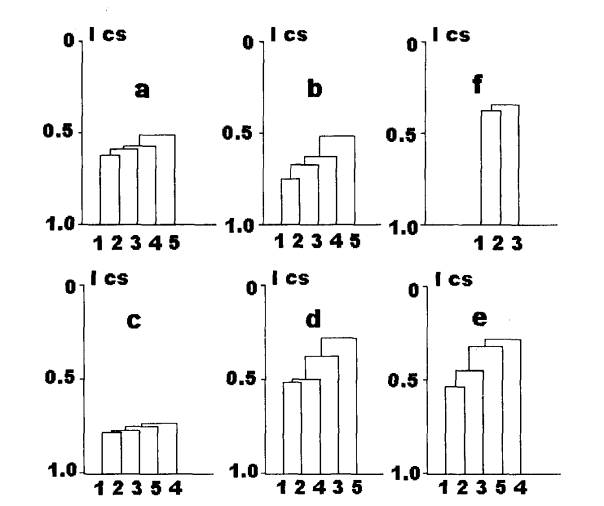
Рис.3. Сходство
паразитофауны рыб разных водоемов бас. Сев.Двины.
a) Сходство паразитофауны рыб разных выделов бас. Сев.Двины.
По оси ординат: индекс Чекановского-Съеренсена (ICS).
По оси абсцисс: 1 — бас. верхней Сухоны, 2 — бас. нижней Сухоны, 3 — бас. р.Вычегда,
4 — бас. оз.Кубенское, 5 — бас. р.Юг.
b)
Сходство паразитофауны рыб разных выделов бас. Сев.Двины (без учета простейших
и моногеней).
Обозначения те же, что на рис. За.
c)
Сходство ихтиопаразитофауны в пределах БР фаунистического комплекса.
Обозначения те же, что на рис. За.
d)
Сходство ихтиопаразитофауны в пределах ПК фаунистического комплекса.
Обозначения те же, что на рис. За.
e)
Сходство ихтиопаразитофауны в пределах АП фаунистического комплекса.
Обозначения те
же, что на рис. За.
О
Сходство ихтиопаразитофауны разных экологических типов водоемов.
По оси ординат:
индекс Чекановского-Съеренсена (ICS).
По оси абсцисс: 1 — крупные реки, 2 — малые реки, 3 — озера.
БП комплекс отдельно не анализируем из-за его неполной изученности
в ряде выделов. Для р.Юг в пределах бас. Сев.Двины, в отличии от р.Вычегды,
характерно наибольшее своеобразие как ПК видов, так и паразитофауны в целом.
Что объясняется, вероятно, не столько их геологическим прошлым, которое у них
было общим (Котласское озеро), сколько
южным географическим положением бас. р.Юг, где лучше смогли сохраниться
теплолюбивые ПК виды и их хозяева. Для своеобразия ихтиопаразитофауны
географическое положение (р.Юг) играет, вероятно, более значимую роль, чем
наличие каналов, соединяющих в настоящее время бассейны Волги и Сев.Двины (через оз.Кубенское и
р.Вычегду). Заметим, что наибольшее сходство фаун всех выделов отмечено для БР
видов, что вообще характерно для этого фаунистического комплекса.
Соотношение различных систематических групп в паразитофауне оказалось
достаточно постоянным для бассейнов рек, оз. Кубенское выделяется большей долей
цестод и трематод (рис.4).
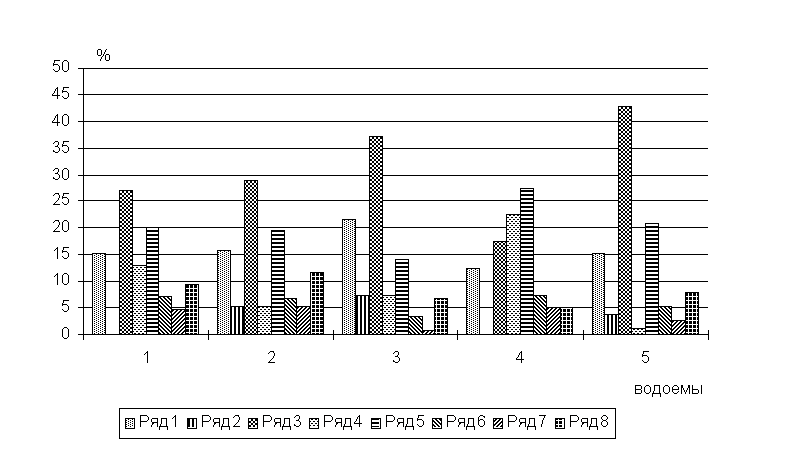
Рис.4. Соотношение основных систематических групп в ихтиопаразитофауне бас. Сев. Двины.
По оси
абсцисс: 1 - бас. верхней Сухоны, 2 - бас. нижней Сухоны, 3 - бас. р.Вычегда, 4
- бас. оз.Кубенское, 5 - бас. р.Юг. Ряды: 1 - миксоспоридии, 2 - инфузории, 3 -
моногенеи, 4 - цестоды, 5 - трематоды, 6 - нематоды, 7 - сребни, 8 -
ракообразные.
Зоогеографические выделы бассейна Северной
Двины, определенные по паразитологическим данным
Помимо примененного выше сравнения водоемов по
фаунистическим комплексам, в зоогеографии
используется количественный подход - сравнение видовых списков территорий
(акваторий) методом кластерного
анализа. Результаты данного метода отчасти наглядно приведены на рис.3. Наибольшее сходство фауны
паразитов наблюдается между верхней и нижней Сухоной, к ним последовательно
примыкают р.Вычегда и оз.Кубенское; р.Юг отличается наибольшим своеобразием
(рис. 3a,b), причины чего обсуждались выше. Указанный порядок сходства сохраняется,
в какой-то мере, и при сравнении видовых списков по отдельно взятым систематическим
группам паразитов. Более точной можно считать дендрограмму, демонстрирующую сходство
только тех групп паразитов, распространение которых в бас. Сев.Двины изучено
достаточно хорошо, а именно всех, кроме простейших и моногеней (рис. 3b). При
исключении из анализа указанных групп, взаиморасположение выделов на дендрограмме
не изменилось, однако, разница в уровнях сходства выделов увеличилась вместе с
ростом индексов сходства (Ics)
(рис. 3a,b).
Паразитофауну рыб бас. Сев.Двины, по сравнению с таковой других
крупных водоемов Восточной Европы, можно считать обедненной. Из числа факторов,
влияющих на богатство фаун (Песенко, 1982), для объяснения обедненности
ихтиопаразитофауны бас. Сев.Двины применимы следующие: 1)временная гипотеза:
более старые сообщества содержат больше видов; 2)гипотеза пространственной
разнородности: в более разнородных физических условиях больше видов; 3)гипотеза
климатической стабильности: в стабильных условиях больше видов.
Наибольшее сходство
паразитофауны рыб среди водоемов, расположенных между водоемами Кольского
полуострова и р.Обь, бас. Сев.Двины имеет с бас.Волги, затем с бассейнами Онежского
и Ладожского озер и с Обь-Иртышским бассейном (Доровских,1990). Нами
установлено, что сходство с бас.Волги оказалось еще выше, чем считалось ранее.
Ихтиопаразитофауна бас. Сев.Двины отличается от паразитофаун других крупных
бассейнов Ледовитоморской провинции наиболее высоким процентом видов
бореального равнинного комплекса (сравнение наших результатов и данных
О.Н.Пугачева (1990).
ЭКОЛОГИЧЕСКИ ОБУСЛОВЛЕННЫЕ РАЗЛИЧИЯ
ПАРАЗИТОФАУНЫ И ИХ СООТНОШЕНИЕ С ГЕОГРАФИЧЕСКИМИ РАЗЛИЧИЯМИ
Как отмечено в научной литературе, при зоогеографическом анализе в
сравниваемых географических выделах одинаково должны быть представлены разные
биотопы (Песенко, 1982). К сожалению, ни в одном из источников использованной
нами литературы по паразитофауне рыб бас. Сев.Двины не оговорено, в какой мере
исследованы разные по экологии водоемы. Вследствие этого, опираясь лишь на наши
данные, рассмотрим отличия паразитофауны разного типа водоемов, расположенных на одной территории. Имеющиеся
материалы позволили выделить лишь три типа водоемов: 1)крупные реки (Сухона,
Юг, Луза); 2)малые реки; 3)озера.
Первая группа водоемов (крупные реки) характеризуется значительными
размерами; наличием течения; меандрирующим руслом, водная растительность
представлена слабо; в ихтиофауне преобладает группа БР видов, значительна доля
ПК видов, из АП видов многочисленен лишь налим, в малых количествах - нельма и
сиг, представители БП комплекса отсутствуют (отмечены только в период покатных
миграций).
Вторая группа водоемов (малые реки), кроме небольших размеров и
наличия течения, особенно сильного на перекатах, отличается достаточно
постоянным руслом, наличием высших водных растений; относительно холодной водой
в летний период (дневные температуры на 2-5°С ниже, чем в других типах
водоемов). Пониженный нагрев воды вызван затененностью малых рек лесом. Наличие
леса по берегам крупных рек, вследствие их значительной ширины, не имеет
большого значения для их прогрева; хотя тот же фактор уменьшает прогрев воды
малых рек, так как узкое русло затеняется лесом достаточно эффективно.
Содержание кислорода в воде малых рек превышает таковое в других типах водоемов
на 0.4-1.2 мг/л. Большая роль высших растений в биотопе должна приводить к
снижению содержания в воде микроэлементов (Золотухин и др., 1995) и биогенов;
воздушные насекомые в летний период составляют одну из преобладающих групп в
питании рыб. Ихтиофауна слагается из представителей БР, БП и АП (налим)
комплексов (наличие последнего не постоянно). Таким образом, по совокупности перечисленных
факторов, малые реки лесной равнины весьма схожи с горными.
Третья группа водоемов - озера, в районе исследований выделяются небольшими
размерами; отсутствием значительной проточности; обилием водной растительности,
илистым дном; вода, как правило, содержит гуминовые вещества, содержание
кислорода в зимний период обычно низкое. Ихтиофауна состоит из БР и ПК видов.
По соотношению доли фаунистических комплексов в паразитофауне
четко выделяются водоемы разных экологических групп (рис.5).
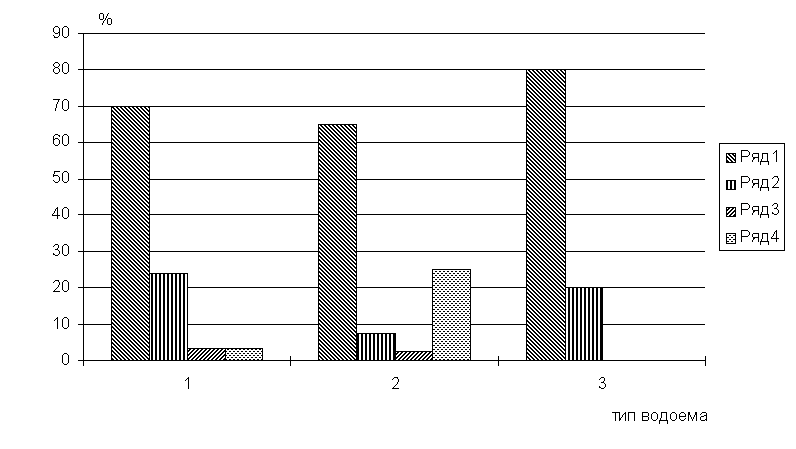 Рис.5. Соотношение доли фаунистических комплексов в
ихтиопаразитофауне разных экологических типов водоемов (бас. Сев.Двины)
Рис.5. Соотношение доли фаунистических комплексов в
ихтиопаразитофауне разных экологических типов водоемов (бас. Сев.Двины)
По оси абсцисс: 1 - крупные реки, 2 - малые реки, 3 - озера. Ряды: 1 - БР виды, 2 - ПК виды, 3 - АП виды, 4 - БП виды
Для крупных рек характерно то же соотношение, что и для паразитофауны
бас.Сев.Двины в целом. Озера отличает отсутствие АП комплекса и БП. Наиболее
своеобразной является группа малых рек, ее характеризует то, что БП комплекс по
числу видов занимает здесь второе место, а не последнее, как в общем
распределении паразитофауны (рис.1). Насколько горная поясность в
распространении ихтиопаразитов аналогична широтной поясности (Шульман, 1958),
настолько же фауна паразитов и рыб, и, вероятно, других гидробионтов малых рек аналогична горной (БП комплекс), и
в меньшей мере - северной (1 вид паразитов АП комплекса). Большое значение для
сходства фауны малых рек равнины с горными реками, как указано выше, имеет
влияние лесного массива. Следовательно, описанная особенность паразитофауны
малых рек характерна лишь для рек лесной зоны. Известно, что в условиях тундры
паразитофауна малых рек, напротив, отличается большей теплолюбивостью,
поскольку малые реки открытых тундровых ландшафтов прогреваются лучше, чем
крупные (Митенев, 1986; 1988).
Кластерный анализ паразитофауны показал, что наиболее сходный
видовой состав паразитов характерен для крупных и малых рек (Ics=0.38)(рис.3f), но даже у
них индекс сходства ниже, чем у самых
отличающихся географических выделов (Ics=0.51-0.52)(рис.3a,b), отсюда можно заключить, что экологические
различия ихтиопаразитофауны в бас.Сев.Двины примерно в 1.5 раза более значимы,
чем географические (при сравнении наиболее сходных пар выделов).
Таким образом, фауна паразитов указанных экологических типов водоемов
представляет собой своего рода эколого-фаунистические комплексы. В таковые
крупных рек (русловой эколого-фаунистический комплекс) входят виды БР - 69.8%,
ПК - 23.8%, АП - 3.2% и БП - 3.2%
фаунистических комплексов. Для озер (озерный эколого-фаунистический комплекс)
отмечены виды БР - 80.0% и ПК - 20.0% фаунистических комплексов, паразитофауна,
в целом, обедненная, но из всех рассматриваемых типов водоемов только в озерах
есть специфичные паразиты карася и красноперки. Для малых рек (притоковый
эколого-фаунистический комплекс) указаны виды БР - 65.0%, БП - 25.0%, ПК - 7.5%
и АП - 2.5% фаунистических комплексов, их отличительной особенностью (малых
рек) является относительно высокая доля БП видов, имеющихся только здесь.
Следовательно, наличие индивидуальности у описанных эколого-фаунистических
комплексов и их конкретная биотопическая приуроченность не вызывают сомнений.
В бас. Сев.Двины зарегистрировано 189 видов паразитов рыб.
В пределах данного бассейна наибольшее сходство видового состава
ихтиопаразитов имеют бассейны верхней и нижней Сухоны, несколько ниже сходство
у них с р.Вычегда, оз.Кубенское и р.Юг. Бассейны оз. Кубенского (и верхней
Сухоны) наиболее отличаются от р.Юг, первый из них выделяется, вероятно, по
причинам исторического плана - длительная изоляция от всей остальной части бас.
Сев.Двины в четвертичном периоде; второй - своим современным географическим
положением (южным).
Расселение фаун в Европейском округе происходило не только в
широтном направлении (восток - запад), как в Сибирском округе (Пугачев, 1990),
но и в меридиональном (из бас. р.Волга), что отразилось на ихтиопаразитофауне:
понто-каспийские (южные) виды распределяются по бас.Сев.Двины по-иному, нежели
все прочие. По сходству фауны ПК видов отчетливо вырисовываются три выдела,
соответствующие истории формирования фауны бас.Сев.Двины: 1) р.Сухона (с
оз.Кубенское), 2) р.Вычегда и 3) р.Юг. Сходство фауны БР видов примерно одинаково
для всех водоемов бас.Сев.Двины, а АП распределены по бассейну в соответствии с
распространением их хозяев.
Из трех экологических типов водоемов (крупные реки, малые реки и
озера) своеобразием ихтиопаразитофауны выделяются малые реки, их
паразитофауна близка к горной;
специфика паразитофауны озер в том, что наряду с обеднением фауны присутствуют
имеющиеся только здесь специфичные паразиты карася и красноперки. Экологические
различия паразитофауны рассматриваемого региона оказались существеннее, чем
географические, на основании чего сделана попытка выделить современные
эколого-фаунистические комплексы паразитов рыб.
Апухтин Н.И., Кузьмин Ю.А., Никонов А.А. и др. Литолого-палеогеографическая карта СССР. Четвертичный период. Время максимального развития позднечетвертичного (калининско-зырянского) оледенения // Атлас литолого-палеогеографических карт СССР. -М.,1967. - Т.4. - С.29.
Берг Л.С. Рыбы пресных вод СССР и сопредельных стран. -М.,1949. - Ч.3. - С.929-1381.
Богданова Е.А., Никольская Н.П. Паразитофауна рыб Волги до зарегулирования стока // Изв. ГосНИОРХ, 1965. - T.60. - С.5-110.
Гнедина М.П., Савина И.В. К фауне паразитических червей рыб Северной Двины. -Вятка,1930. - С.87-106.
Гусев А.В. Моногенетические сосальщики рыб системы реки Амур // Труды зоол. ин-та АН СССР, 1955. - T.19. - С.171-398.
Догель В.А. Значение паразитологических данных для решения зоогеографических вопросов // Зоол. журнал, 1947. - T.25. -Вып.6. - C.481-492.
Доровских Г.Н. Паразитофауна рыб среднего течения реки Вычегды: Автореф. на соиск. уч. степ. канд. биол. наук. - Л., 1988. - 25 c.
Доровских Г.Н. Зоогеографическая характеристика и история формирования паразитофауны рыб Северо-Двинского бассейна //Биологические ресурсы Белого моря и внутренних водоемов Европейского Севера. - Сыктывкар, 1990. - С.22.
Екимова И.В. Эколого-географический анализ паразитов рыб реки Печоры // Болезни и паразиты рыб Ледовитоморской провинции. - Свердловск, 1976. - С.50-68.
Зверева О.С. Особенности биологии главных рек Коми АССР. - Л., 1969. - 279 c.
Золотухин И.А., Никулина С.Н., Федосеева Л.А. Снижение концентрации микроэлементов в водной среде под воздействием корневых систем // Экология, 1995. - №3. - С.128-249.
Ивашевский Г.А., Доровских Г.Н. Паразитофауна гольяна обыкновенного реки Шарденьга (бассейн р. Северной Двины) // Материалы II молодеж. науч. конференции: “Рациональное использование и воспроизводство природных ресурсов”. - Сыктывкар, 1993. - С.20-21.
Ивашевский Г.А. Видовой состав паразитов некоторых видов рыб бассейна нижнего течения р. Сухона // Тезисы региональной студ. науч. конференции: ”Европейский Север в вузовском краеведении”. - Вологда, 1994. - С.51-52.
Ивашевский Г.А. Видовой состав паразитов некоторых видов рыб бассейна верховьев Северной Двины // Тезисы международ. науч. конференции: “Биологические ресурсы Белого моря и внутренних водоемов Европейского Севера”. - Петрозаводск, 1995. -С.93-95.
Ивашевский Г.А. Первые данные по паразитофауне рыб р.Луза// IV молодеж. науч. конференция Ин-та биол. Коми науч. центра УрО РАН: “Актуальные проблемы биологии”. - Сыктывкар, 1996. - С.53.
Квасов Д.Д. Позднечетвертичная история крупных озер и внутренних морей Восточной Европы. - Л.,1975. - 278 c.
Кудрявцева Е.С. Паразитофауна рыб р.Сухоны и Кубенского озера // Зоол. ж., 1957. - T.36. - Вып.9. - С.1292-1301.
Кудрявцева Е.С. Фаунистический обзор паразитов рыб р.Сухоны и Кубенского озера: Сообщение 1-е // Уч. зап. Вологодского гос. пед. ин-та, 1959. - T.24. - С.175-185.
Кудрявцева Е.С. Фаунистический обзор паразитов рыб р.Сухоны и Кубенского озера: Сообщение 2-е // Там же. 1962. - T.27. - С.219-254.
Кудрявцева Е.С. Фаунистический обзор паразитов рыб р.Сухоны и Кубенского озера: Сообщение 3-е // Там же. 1966. - T.29. - С.191-201.
Митенев В.К. Миксоспоридии рыб Кольского полуострова // Морфология, систематика и фаунистика паразитических животных: Труды зоол. ин-та АН СССР. - Л., 1986 - Т.155. - С.134-154.
Митенев В.К., Шульман Б.С. Моногенеи рыб Кольской субарктики // Исследования моногеней: Труды зоол. ин-та АН СССР. -Л.,1988. - Т.177. - С.60-76.
Определитель паразитов пресноводных рыб фауны СССР. - Л.,1984. - Т.1. - 428 с.; - Л.,1985. - Т.2. - 425 с.; - Л.,1987. - Т.3. - 583 с.
Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. - М., 1982. - 284 с.
Пугачев О.Н. Моногенеи пресноводных рыб северо-востока Азии// Исследования по морфологии и фаунистике паразитических червей: Труды зоол. ин-та АН СССР. - Л.,1983. - Т.121. - С.22-34.
Пугачев О.Н. Гельминты пресноводных рыб северо-востока Азии // Там же. - С.90-113.
Пугачев О.Н. Зоогеографические особенности паразитофауны рыб Ледовитоморской провинции // Паразиты и болезни гидробионтов Ледовитоморской провинции. - Новосибирск, 1990. - С.5-15.
Хотеновский И.А. Фауна СССР. Моногенеи. - Л., 1985. - 262 c.
Шульман С.С., Шульман-Альбова Р.Е. Паразиты рыб Белого моря. - Л., 1953. - 198 c.
Шульман С.С. Зоогеографический анализ паразитов пресно-водных рыб Советского Союза // Основные проблемы паразитологии рыб. - Л.,1958. - С.184-230.
Яковлев В.Н. Распространение пресноводных рыб Неогена в Голарктике // Вопросы ихтиологии, 1961. - Т.1. - Вып.2. - С.209-220.
ОГЛАВЛЕНИЕ
Введение................................................ 3
Материал и методика..................................... 4
Зоогеографический анализ ихтиопаразитов бассейна Северной
Двины................................................... 5
Фаунистические
комплексы в паразитофауне
рыб бассейна Северной Двины......................... 5
Зоогеографические выделы бассейна Северной
Двины, определенные по паразитологическим данным..... 11
Экологически обусловленные различия паразитофауны и
их соотношение с географическими различиями ........... 13
Заключение.............................................. 16
Литература.............................................. 17